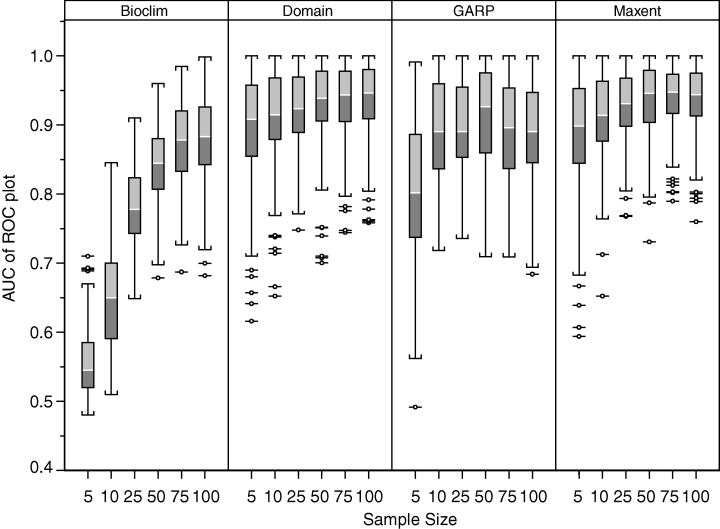
Species, especially those of interest to conservationists, often have limited to rare occurrence data. This “sparseness” of the occurrence data poses problems with developing accurate distribution models/predictions. Previous studies have examined the effect of sample size on the accuracy of these SDMs; however, the sample size of individual species has not been investigated. Individual species characteristics, such as range size, may impact the accuracy of the models. The authors examine the impact of sample size on model accuracy for 18 taxa (California taxa) using 4 modeling methods using presence only data.
Of the methods used, MaxEnt provided the most useful results even as small sample sizes (5, 10, 25, of the maximum 150). The Domain and GARP performed reasonably well with the small sample sizes. However, Bioclim performed with worst. They also point out that multiple measures of model accuracy, not just AUC, are needed to determine the performance. In terms of species characteristics on model performance, species with smaller ranges, both geographical and environmental, provided greater accuracy in the models. The authors then highlight that these models should be used by conservationists to estimate rare species distributions.
Hernández, P. A., Graham, C. H., Master, L. L. & Albert, D. L. 2006 The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography (Cop.). 29, 773–785. (doi:DOI 10.1111/j.0906-7590.2006.04700.x)
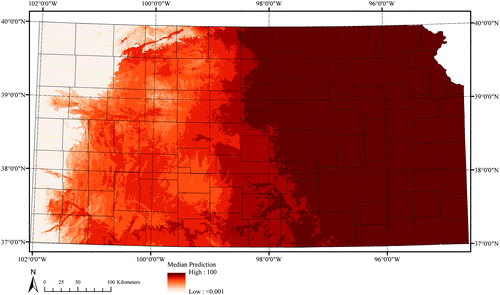
 Female adult lone star tick (Amblyomma americanum)
Female adult lone star tick (Amblyomma americanum)