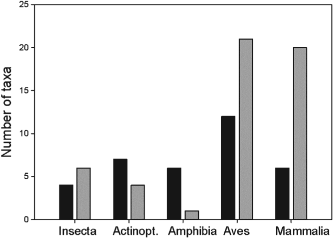
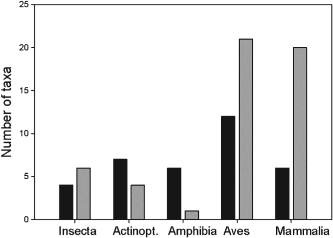
Integrating biotic interactions into the framework of species richness models has been a suggested to improve the performance of both species distribution models. The authors seek to use biotic variables in two species richness modeling frameworks. Stacked species distribution models (SSDM) fit separate species distribution models then blindly stacks the results of the predicted occurrences to calculate the species richness. The macroecological models (MEM) do not use the information provided by species identity and community composition to estimate the species number using environmental conditions. This model assumes species richness is limited by environmental conditions. Using these two models, three different groups of taxa (vascular plants, bryophytes, and lichens) used to examine the effect of integrating biotic variables.
When comparing the results of the models using biotic interactions to models with only climatic and abiotic, biotic models performed consistently better. Both modeling frameworks and all taxonomic groups using biotic interactions had a lower bias and increased predictive power. These results highlight the importance of using biotic predictor variables in not only species richness models but also single species distribution models.
Mod, H. K., le Roux, P. C., Guisan, A. & Luoto, M. 2015 Biotic interactions boost spatial models of species richness. Ecography (Cop.). 38, 913–921.

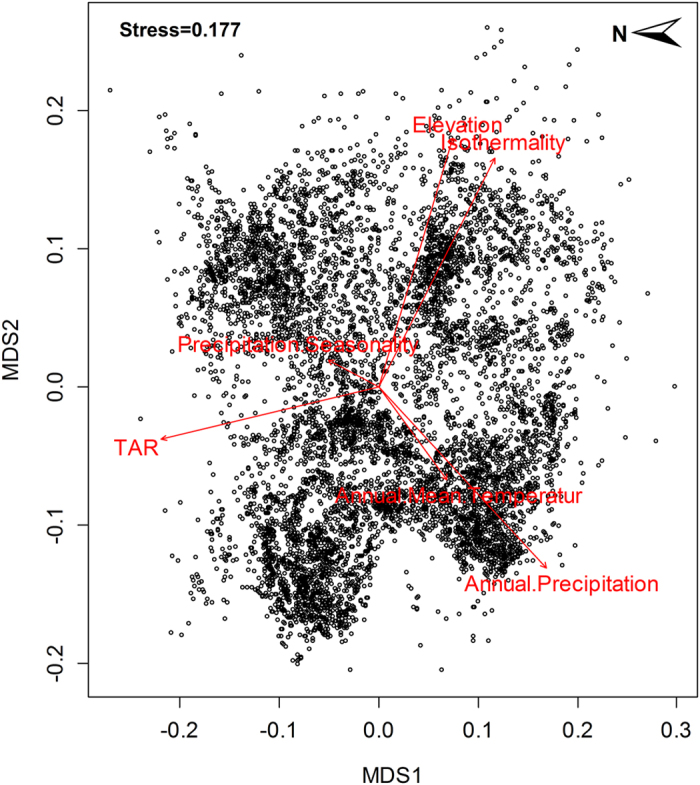
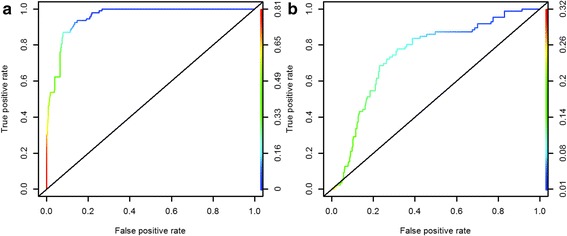
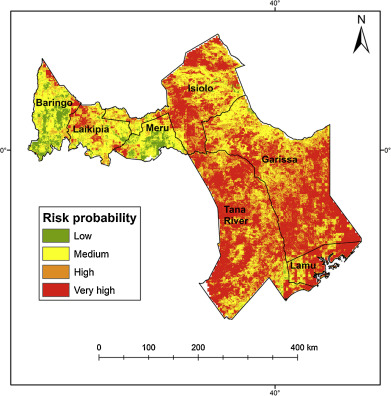
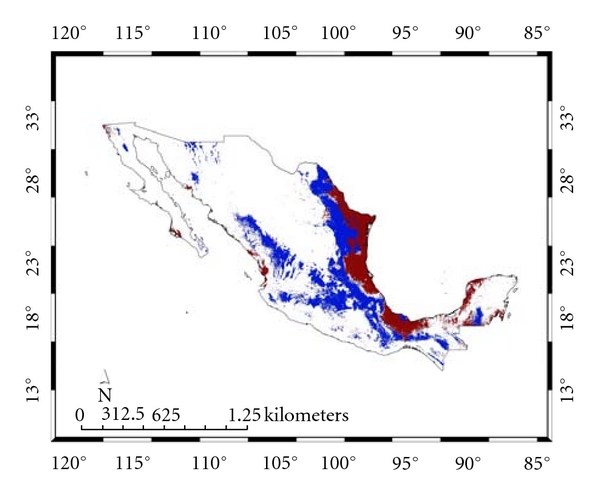

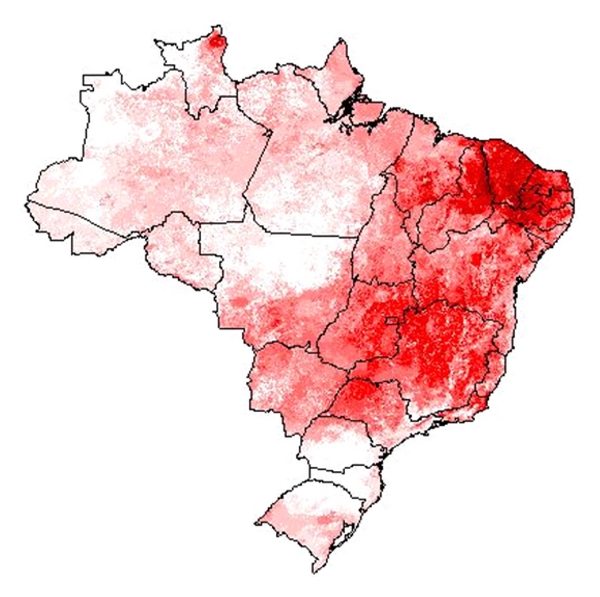

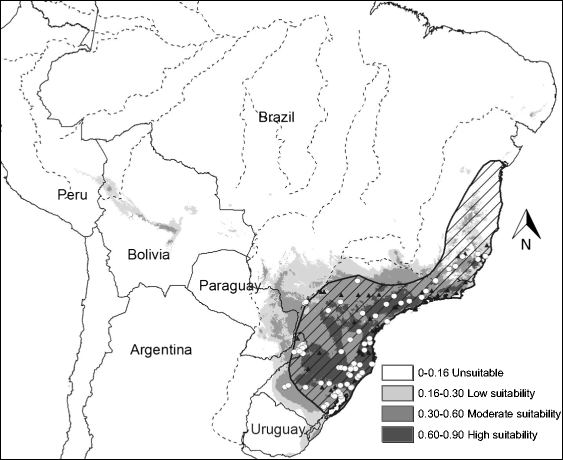
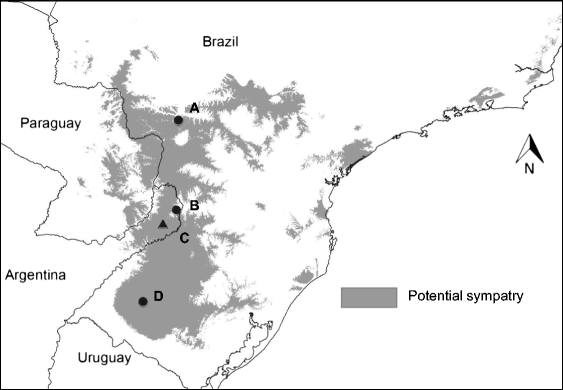
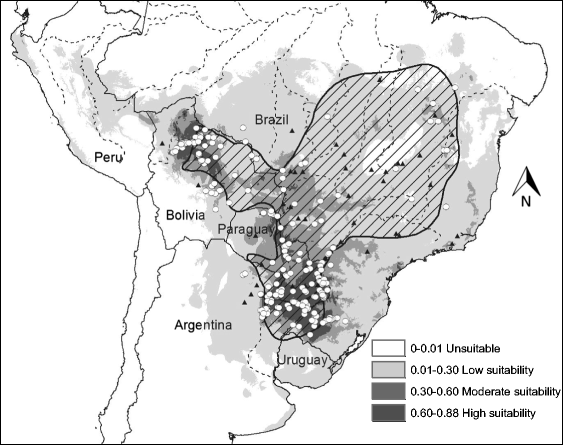 Habitat suitability (low, moderate, high) categorized the resulted of the predicted areas. In addition, the models were evaluated using AUC. Both the models provided a wider distribution than currently estimated. The results of the model for Alouatta caraya had a broader range and a variety of temperatures with Temperature Annual Range as the most influential to the distribution.
Habitat suitability (low, moderate, high) categorized the resulted of the predicted areas. In addition, the models were evaluated using AUC. Both the models provided a wider distribution than currently estimated. The results of the model for Alouatta caraya had a broader range and a variety of temperatures with Temperature Annual Range as the most influential to the distribution.