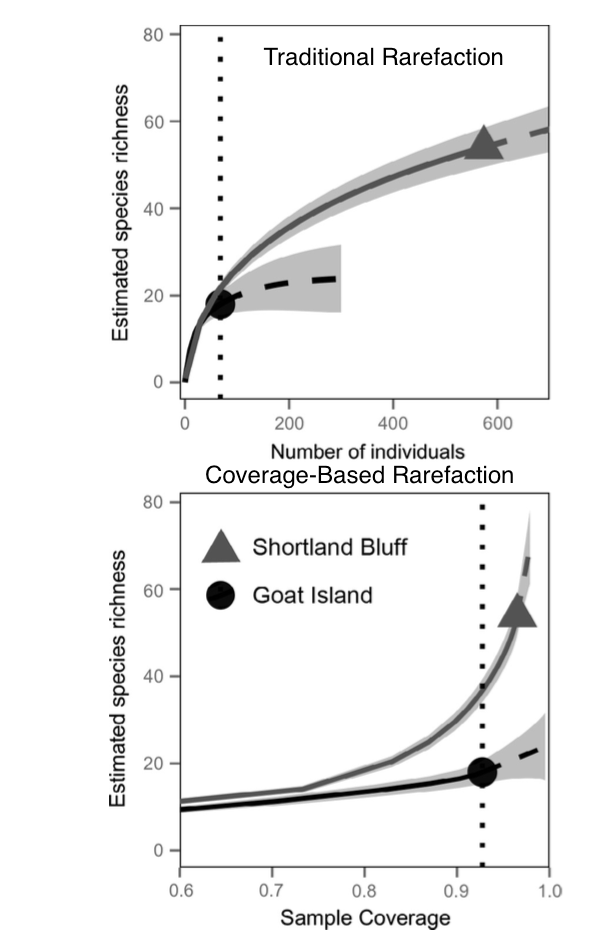
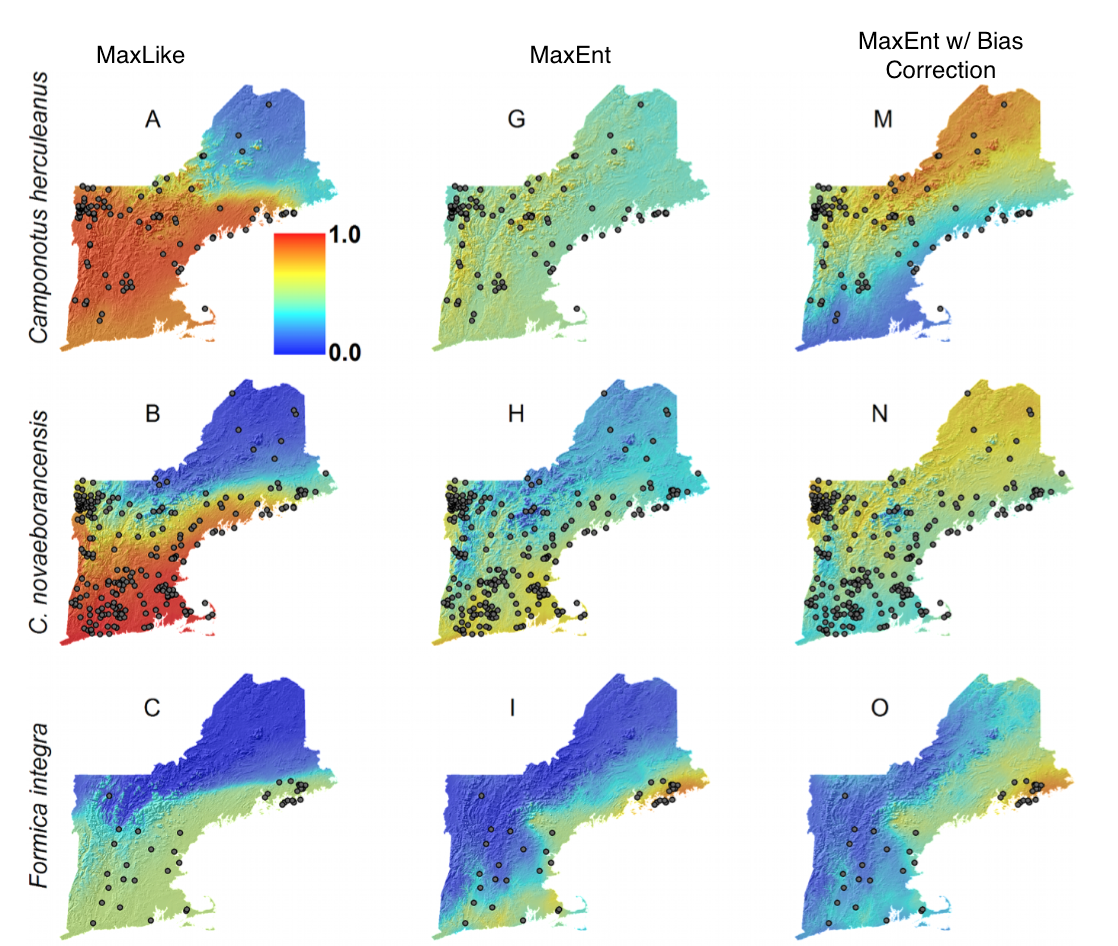
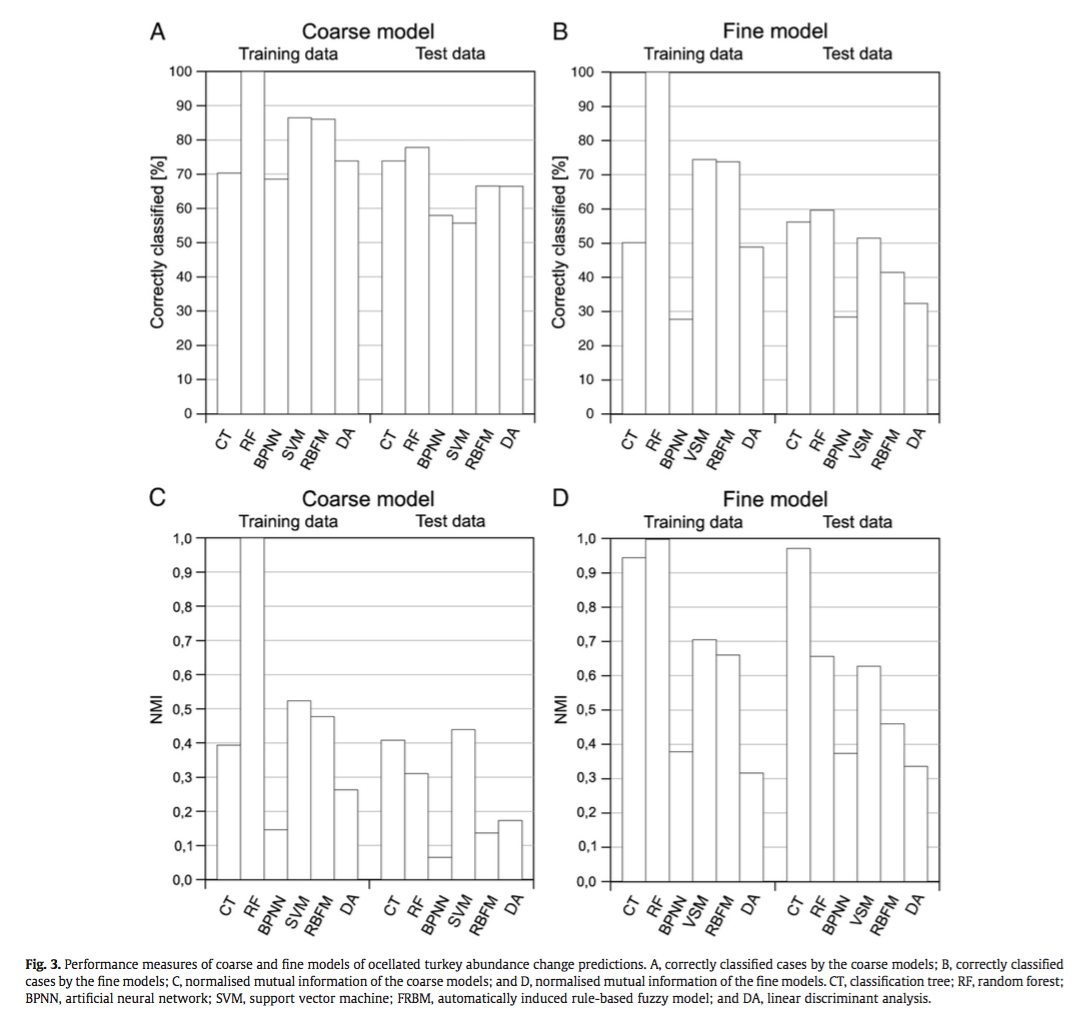
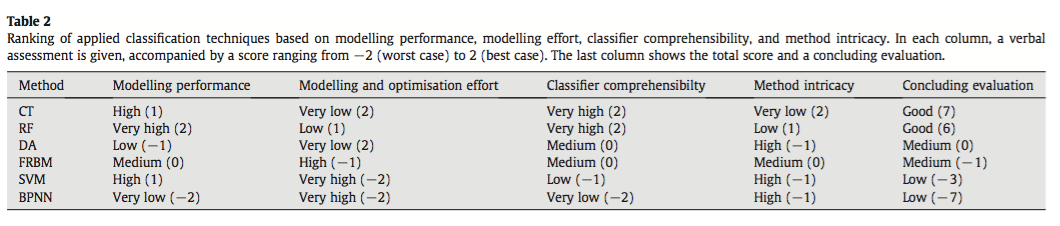
Liu, C., White, M., & Newell, G. (2013). Selecting thresholds for the prediction of species occurrence with presence-only data. Journal of Biogeography, 40(4), 778–789. http://doi.org/10.1111/jbi.12058
Many of the newer methods for SDM output continuous values, but binary predictions are often needed in application and when evaluating models, necessitating the need for thresholding. Many thresholding techniques have been proposed, such as lowest presence threshold, fixed thresholding, and minimizing or maximizing a the difference between opposing statistics, but there is no consensus on which performs better, and presence-only data can not be used in all of these instances. Liu et al (2013) compared 12 different techniques through mathematical proofs and simulations of species distributions. First, they use mathematical proofs to show that only eight of the techniques will result in similar thresholds using presence-only and presence-absence (or pseudo-absence) data. They then use 1000 simulated data sets of species occurrences to evaluate the variation within each thresholding technique, based on eight modeling approaches: Mahalanobis distance, ecological niche factor analysis, and GAM and random forest, the final two which each had three models using presence/absence, presence/pseudo-absence, and presence/psuedo-absence filtered with Mahalanobis distance. They choose four techniques (max kappa, min D01, meanPred, and max SSS) to evaluate. To sum up their (many) results, max SSS (which maximizes the sum of sensitivity and specificity) was most robust to pseudo-absences and changes in species prevalence, meaning it had less variation in its threshold definition as those parameters changed. When evaluated on the criteria of objectivity, equality, and discriminability, it performed better than the other methods because it is objective, unaffected by pseudo-absences, and produced higher sensitivity and specificity than other methods. All of the thresholding methods, however, are influenced by sampling bias in environmental space, especially as the sample size decreases. Based on all of the caveats to thresholding, I think it would be best to avoid doing unless absolutely necessary or to offer results based on multiple thresholds. When this is unavoidable, max SSS will perform the best.