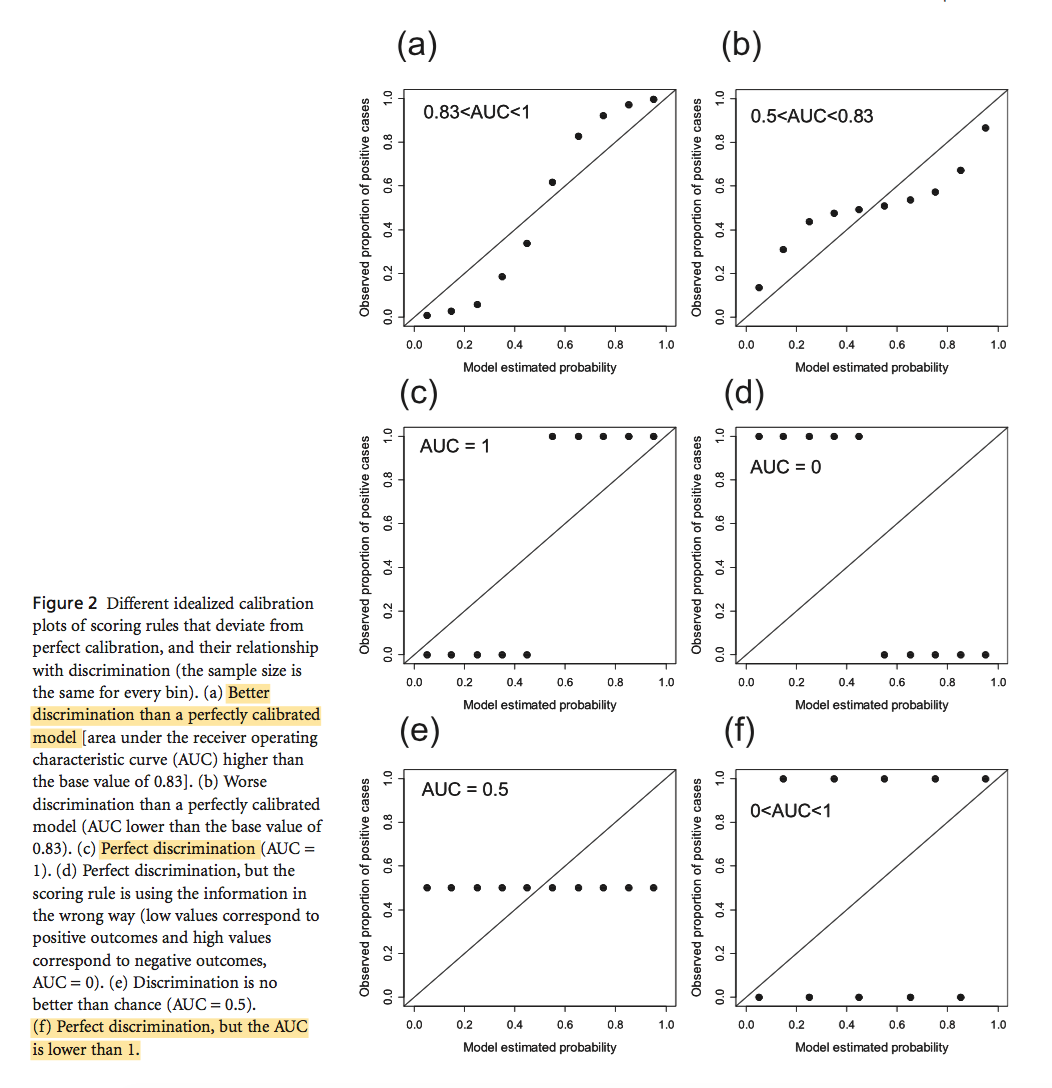
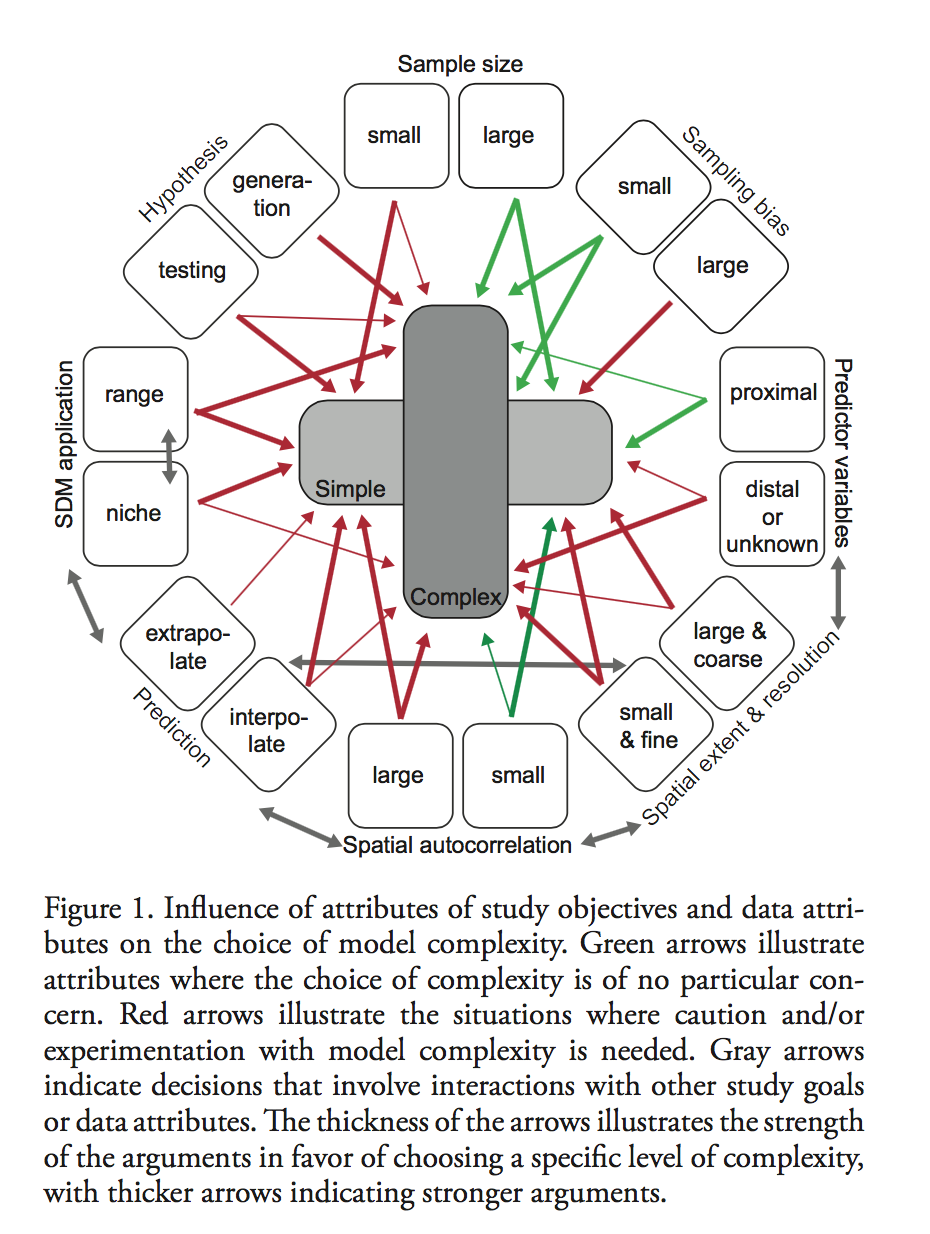
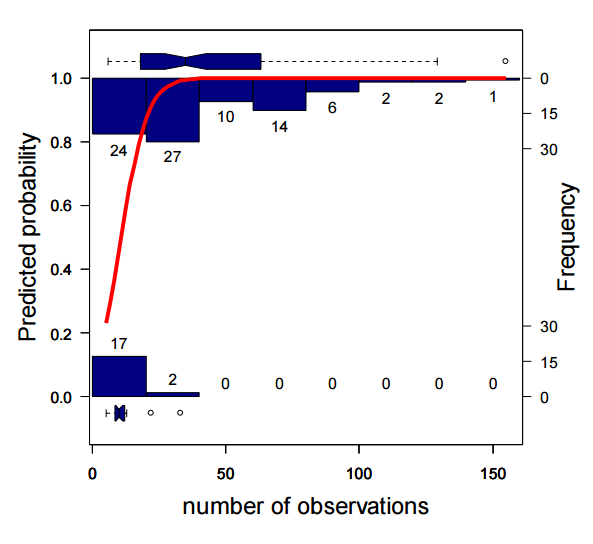
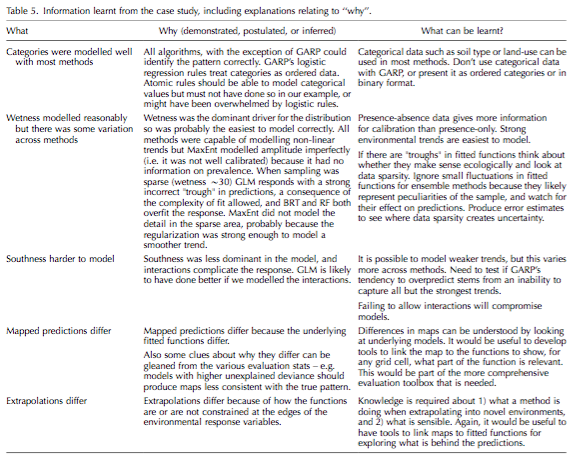
The vulnerability of species to range expansion by predators can be predicted using historical species associations and body size. Declines in abundance in local extinctions are the direct consequence of climate exceeding physiological tolerances in addition to the indirect consequences of climate change on species interaction. These indirect impacts of climate change and biodiversity are more difficult to predict or observed when compared to the physiological impacts.
Species ranges have changed at variable rates under climate change, potentially making novel ecosystems. However, species expanding their range can encounter resident prey, predators and competitors that were present in their historical range (i.e. species historically occurred in some patches). The ecological niche concept has been used to understand patterns of co-occurrence and species interaction; this could also be a useful tool to protect the indirect impacts of climate change.
Here, the authors suggest using species associations and body size as a simple measure of the impacts of species introductions facilitated by climate change. Negative associations can indicate strong ecological interactions including competitive exclusion, predation or it could indicate different abiotic requirements. Functional traits often mediate the strength of species interactions – which can be used to infer niche differences. Body size is correlated with many functional traits (i.e. reproductive rate, dispersal ability, diet breadth and or predation). Increased differences in body sizes would indicate decreased competition, while the ratio of predator to prey body size indicates strength of predation.
The authors hypothesize that pairwise species associations and body size can predict the relative risk imposed on resident species by predators whose ranges are expanding. They focus centrarchid predators undergoing range expansion in the Great Lakes region. This expansion is expected to be problematic since these predators are not often found in smaller lakes with the potential prey species. The question then becomes whether this negative species associations are good predictors of vulnerability, and how resident species body size impacts the risk associated with additional predators.
Methods: The data set consisted of 1551 links with paired historical and contemporary species samplings. A total of 106 fish species were observed which was then used to create presence absence data pairs in 2 x 2 contingency tables. The Phi– coefficient was calculated for these 2 x 2 tables (range from -1, 1) the relative risk ratio was then calculated on the tally of lakes where the resident species was absence after the introduction of the predator.
Results: Centrarchid introductions significantly increase the likelihood of some prey species loss, while protecting loss of native centrarchids based on introduction data. Historical species associations were a strong predictor of the introduced species’ impact. Additionally, resident species total length was a significant indicator of the relative risk ratio.
Take home: Traits mediate species interactions, and body size is an easily measurable trait that is correlated to many other traits in fish species. Body length and historical species associations can be used to forecast the impact of introduced species on the native species under climate change.
Given that fish can have convoluted food webs, using body size as a proxy of competition and predation seems like a very elegant solution.
Alofs, Karen M., and Donald A. Jackson. “The vulnerability of species to range expansions by predators can be predicted using historical species associations and body size.” Proc. R. Soc. B. Vol. 282. No. 1812. The Royal Society, 2015. http://dx.doi.org/10.1098/rspb.2015.1211