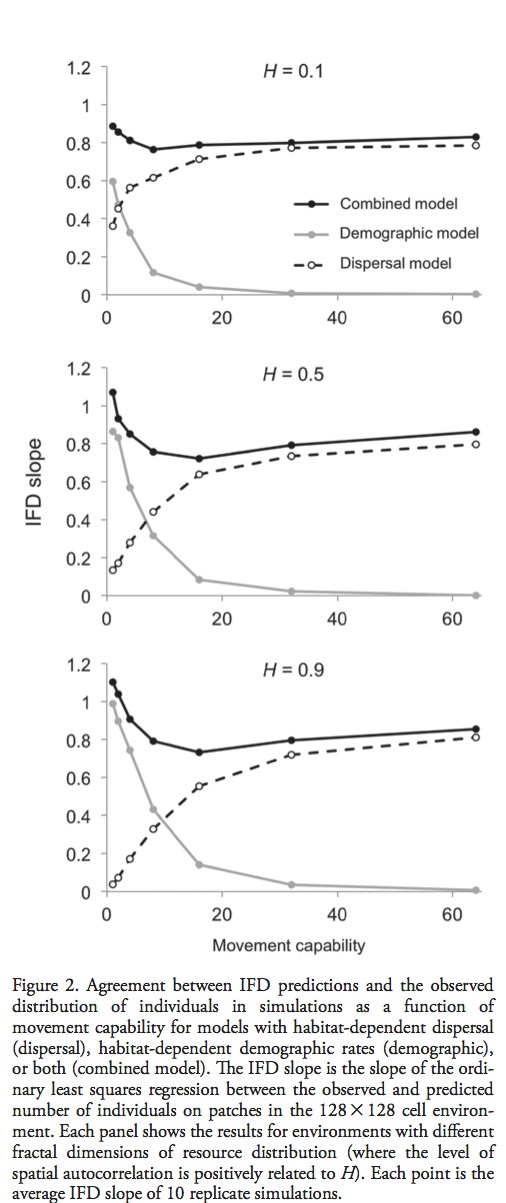
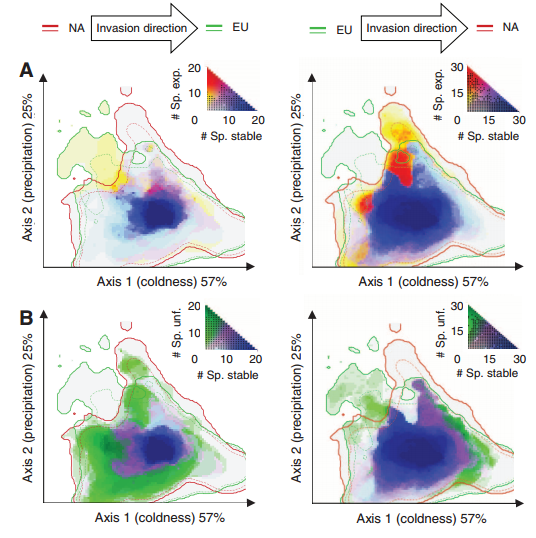
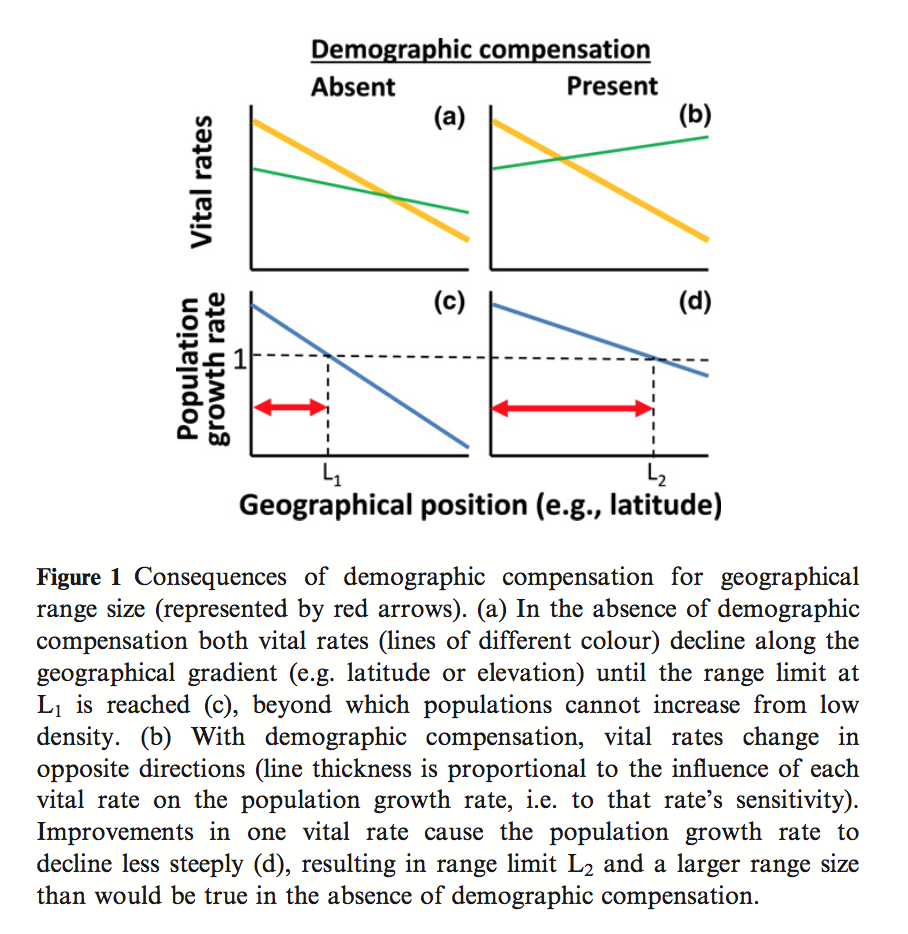
These papers discuss and argue the terminology used to describe the process of determine where species are able to inhabit. Specifically the use of the terms Ecological Niche Models and Species Distribution Models to describe the techniques used to determine where or potentially where species are able to inhabit.
Warren (2012) criticizes the “loss” of the niche in the terminology. While he sympathizes that many of these models are trained using data that only comes from the distribution of a species, he also argues that the underlying assumption of these models is that they are estimating the niche. The argument that these environmental predictors have some effect on biological processes of the organism and that often these models omit processes (e.g. dispersal). Here, he defines the niche as the “conditions within which the species can survive and reproduce”. He suggests we continue to acknowledge the conceptual framework being used and the we are attempting to estimate the niche in our research.
In a response to Warren, McInerny and Ettienne (2013) defend the position of using ‘distribution’ to describe the techniques. They criticize his definition (even the definitions he provides, pointing out that he invokes two ones) of niche saying it constrains the selection of predictor variables to ones that are only biological. However, they point out that these problems have also been considered with the SDMs through parameter selection, model structure, and functional forms. Lastly, the point out that other words could also be used to describe these models (“habitat suitability”, “bioclimate envelopes”, “resource selection”) but they stand by the choice of the neutral words “species distribution modeling”.
Warren, D. L. 2012 In defense of ‘niche modeling’. Trends Ecol. Evol. 27, 497–500. doi:10.1016/j.tree.2012.03.010
McInerny, G. J. & Etienne, R. S. 2013 ‘Niche’ or ‘distribution’modelling? A response to Warren. Trends Ecol. Evol. 28, 191–192. http://dx.doi.org/10.1016/j.tree.2013.01.007